|
|
|
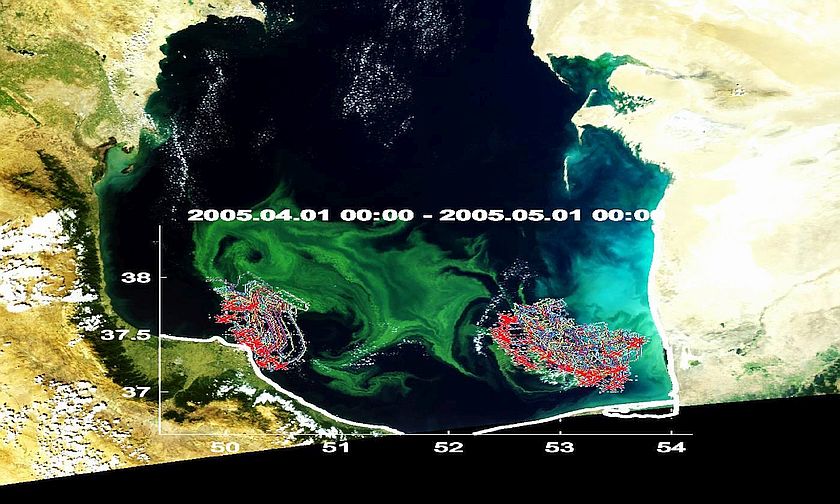
Введение. Каспийское море, в соответствии с физико-географическими и геолого-геоморфологическими признаками, подразделяют на три части: Северный Каспий, Средний и Южный. Первой характерной особенностью Северного Каспия является расположение в пределах шельфовой зоны, что обуславливает малые глубины (преимущественно до 5 м, максимальные – не более 15-20 м) водоема. Вторая особенность связана с поступлением большого количества пресных вод: более 80% общего стока вносится реками (в основном – Волгой) в северную часть моря [11]. Южный Каспий, напротив, отличается максимальными глубинами (Южнокаспийская котловина – 1025 м [5]) и незначительным вкладом речных стоков (около 3-5% от общего стока [10]). При исследовании цветений сине-зелёных водорослей в границах водосборного бассейна Каспийского моря немалое внимание уделялось распространению этого вида фитопланктона по руслу Волги. Так, указывалось, что в водохранилищах Волжского каскада [6] высокое содержание биогенных элементов является благоприятным условием для развития цианобактерий. Там же подчеркивается проблема вторичного загрязнения с негативными проявлениями в виде заморных явлений и ухудшения санитарного состояния водоемов. При исследовании фитопланктона в Куйбышевском водохранилище, также входящем в речную систему Волги [20], авторами были отмечены аномальные цветения сине-зелёных водорослей в 1989 и 1990 гг. При анализе факторов, способствующих росту цианобактерий, не обнаружилось существенное влияние гидрометеорологических условий на изменения концентрации хлорофилла-а, но при этом отмечалась некоторая зависимость от температуры воды и ветровых характеристик. В то же время акцент был сделан на связь цветения с уровнем воды: при снижении уровня риск цветения сине-зелёных водорослей возрастает. Развитие фитопланктона (различный видовой состав) преимущественно в мелководных районах описывалось по отношению к ряду других акваторий – Японскому морю [15], Черному [16], Баренцеву [13], Беринговому [19] и другим. В 2009 г был совершен рейс на юге Каспийского моря, во время которого выполнялся отбор проб [23], и по результатам их анализа было получено, что фитопланктон группы Cyanophyta составлял 98 и 96% численности и биомассы фитопланктона соответственно. В группе Cyanophyta выделились два вида: Oscillatoria sp. и N. Spumigena, причем, последний имел наибольшую численность и массу – более 99%. В настоящее время N. Spumigena (Nodularia Spumigena) оценивается как вид, который развивается в солоноватых водах [22] и выделяет нодулярин, вызывающий гепатоксическое действие. Это биохимическое соединение оказывает отравляющий эффект на многие организмы водной экосистемы (включая беспозвоночных и рыб), при употреблении зараженной воды – на теплокровных, а также накапливается в тканях моллюсков [25]. Учитывая характерные особенности развития сине-зелёных водорослей в общем и N. Spumigena в частности, вполне объяснимо выраженное развитие этого вида фитопланктона в северной части Каспийского моря, где соленость не превышает 10-11‰, глубины – 20 м, и имеется мощный речной сток, уже сам содержащий не только биогенные элементы, но и цианобактерии [18]. Каковы же причины и фоновые условия для возникновения экстремального роста биомассы сине-зелёных водорослей в Южном Каспии, для которого характерны глубоководность, несколько более высокая соленость (до 13‰) [14] и слабо развитая речная сеть? Методы и материалы. Для характеристики случаев аномального цветения был рассмотрен период с 2001 по 2017 г. (с мая по сентябрь). Случаи аномального цветения выявлялись по данным дистанционного зондирования двумя способами: учитывались изменения концентрации хлорофилла-а и изменения яркости восходящего излучения в оптическом диапазоне. В первом случае привлекались данные сенсоров SeaWIFS (1997-2010 гг., временная дискретность – 30 дней) и MODIS Aqua (2002-2017 гг., временная дискретность – 8 дней), по результатам анализа которых было выявлено экстремальное увеличение биомассы фитопланктона в 2001 и 2010 гг. Во втором случае использовались спутниковые снимки среднего (SeaWIFS, MODIA Aqua, MODIS Terra, NOAA-17) и высокого (Landsat-7, Landsat-8, Sentinel-2) разрешения. Их данные позволили обнаружить аномальное цветение характерного для акватории фитопланктона (точный видовой состав не известен, основу могли составлять диатомовые, динофитовые, зелёные и др. [3, 7, 8, 9, 12]) также в 2001 и 2010 гг., и сине-зелёных водорослей в 2005, 2008, 2009, 2010, и 2017 гг. Кроме того, производилась оценка температуры поверхности Каспия по ежесуточным данным: MODIS Aqua (MODISA_L3m_SST) – Sea Surface Temperature at 11 microns (Day or night), GHRSST Level 4 MUR Global Foundation Sea Surface Temperature Analysis (v4.1), NOAA-17 (Environmental Data from Polar-orbiting Satellites (Advanced Very High Resolution Radiometer (AVHRR)). Характеристика синоптической обстановки над южной частью Каспийского моря воспроизводилась по данным: о полях скорости ветра на высоте 10 метров National Centers for Environmental Prediction (NCEP) с 6-часовой дискретностью; NOAA Operational Model Archive Distribution System (NOMADS) с 6-часовой дискретностью; двухмерной модели MERRA Model: Wind speed at 10 m above displacement height, time average (MAT1NXSLV v5.2.0) с часовой дискретностью; модели реанализа MERRA-2 Model: Surface wind speed (M2IMNXLFO v5.12.4) и Surface wind speed, Instantaneous (M2I1NXLFO v5.12.4) с месячной дискретностью. Выполнялась оценка распространения речных вод, для чего привлекалась разработанная в Морском гидрофизическом институте модель воспроизведения траектории плавающих объектов FOTS, которая при выполнении расчетов обращается к архиву данных AVISO и данным о параметрах ветра NCEP и NOMADS. Дополнительно рассматривались пыльные бури, как возможный фактор, способствующий возникновению и развитию аномальных цветений – их частота и интенсивность оценивались по ежесуточным спутниковым снимкам среднего разрешения (Suomi NPP, MODIS Aqua и Terra), изменениям оптической толщины аэрозоля (Aerosol Optical Depth 670 nm (Ocean-only) (SWDB_L3M05 v004)) и вариациям количества (веса) аэрозольных частиц (MERRA-2 Model – Dust Dry Deposition Bin-4 (M2TMNXADG v5.12.4). Обсуждение результатов. За период с 2001 по 2017 гг. в южной части Каспийского моря было выявлено два типа аномального цветения – в 2001 и 2010 гг. цветение типичного фитопланктона и цветение сине-зелёных водорослей (2005, 2008, 2009, 2010 и 2017 гг.). Причиной возникновения экстремального цветения в 2001 г считается занесение в акваторию моря гребневика Mnemiopsis [4]. Наиболее выраженный рост биомассы фитопланктона по данным сенсоров SeaWIFS и MODIS Terra прослеживается с 14 июля по 23-24 августа, но видимом диапазоне присутствие фитопланктона определяется уже с последней трети апреля. Весной водоросли наиболее сконцентрированы в южной половине рассматриваемой части моря, к июню также выделяются на больших площадях вдоль западного побережья. Тем не менее, при массовом развитии в пространственном отношении, количество биомассы фитопланктона не выходит за типичные значения, наблюдаемые в иные года. Первые признаки интенсивного развития цветения наблюдаются с 12-13 июля, характеризуются резким возникновением. Цветение проявляется вдоль практически всего восточного шельфа, на непродолжительный период (с 14 по 16 июля) формируется малая по площади зона повышенной концентрации биомассы фитопланктона, которая в силу недостаточности данных не поддаются видовой идентификации. Аналогичный случай регистрируется позже – с 6 по 16 августа возникает малая область с экстремально высоким (по сравнению с окружающими водами) содержанием фитопланктона, которая с 12 августа разделяется на две части. По косвенным признакам (интенсивность цветения по всей южной части моря в видимом диапазоне, концентрация хлорофилла-а) допустимо предположить, что эти случаи не являются цветением сине-зелёных водорослей. Вне описанных ситуаций аномальное цветение характерного для акватории фитопланктона к 19 июля распространяется вдоль восточного побережья от южного берега до полуострова Челекен и севернее, одновременно захватывая воды в западном направлении. Примерно в этот же период динамичного развития (с 12-13 июля) происходит рост биомассы вдоль южного берега и частично – западного. Дни примерно с 22-23 июля до 5-6 августа можно охарактеризовать как первый пик максимально развитого цветения (рис. 1): захвачены воды всей акватории, наиболее выражено – юго-восточной половины Южного Каспия, отчетливо выделяются суб- и мезомасштабные процессы (фитопланктон выполняет функцию трассера). Второй пик имеет меньшую продолжительность – примерно с 13 по 15-16 августа, области с наиболее высоким содержанием биомассы фитопланктона (на RGB-композитах в псевдонатуральных цветах эти области выглядят зелёными) занимают порядка 70% Южного Каспия.
В 2010 г. общая продолжительность экстремального цветения типичного фитопланктона составила около 5 недель (с 8 мая по 10-12 июня). Возникновение, как и в 2001 г, было резким, но развитие началось с западного и юго-западного побережья с распространением практически на всю южную часть Каспийского моря (на пике – до 75-80% захваченных вод). Тем не менее, по сравнению с летом 2001 г. интенсивность цветения была существенно ниже – максимальные значения концентрации хлорофилла-а (по данным SeaWIFS с 8-дневной дискретностью на 9-километровой сетке) в 2001 г. достигли 15,8 мг/м³, тогда как в мае 2010 г. – только 2,5 мг/м³. Каждый случай цветения сине-зелёных водорослей имел индивидуальную продолжительность – от нескольких дней до нескольких недель, при наблюдении за эволюцией аномальных цветений цианобактерий средствами дистанционного зондирования обычно выделялись достаточно выраженные этапы развития, которые отображены в таблице ниже (табл. 1).
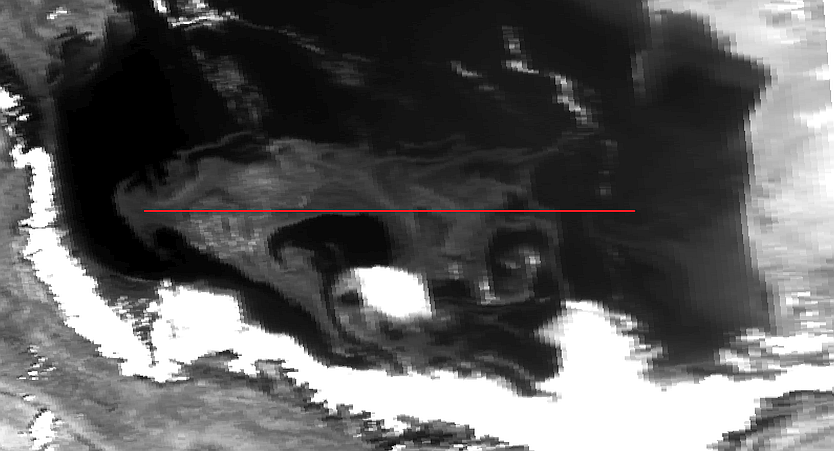
Фаза начального развития характеризовалась ростом биомассы фитопланктона в подповерхностном слое без характерных признаков преобладания цианобактерий; нередко можно было выделить увеличивающуюся в масштабах структуру, сформированную динамическими процессами (течениями, вихрями). Во время фазы максимального развития биомасса фитопланктона достигала максимального объема (на основе оптических характеристик), происходил подъем на нулевой горизонт с последующим развитием на поверхности (проявлялось свойство положительной плавучести), присутствовали характерные признаки для сине-зелёных водорослей (значения отраженной компоненты, нитевидная структура на оптических изображениях высокого разрешения). Фаза регрессии отмечалась противоположными симптомами: снижалось количество биомассы цианобактерий, уменьшалась площадь захваченных вод, на серии спутниковых снимков наблюдалось заглубление и/или исчезновение концентрированных образований экстремального цветения. Допускается, что одним из факторов, способных оказывать влияние на возникновение и дальнейшее формирование аномальных цветений, является температура деятельного слоя акватории [2, 21]. Для южной части Каспийского моря межгодовая изменчивость температуры поверхностного слоя вод характеризуется следующим образом: минимальные значения (по данным ИЗС MODIS Aqua) регистрируются в феврале, затем начинается прогрев, который достигает максимума в августе (до 30°С и более), а с сентября следует период охлаждения. Но при индивидуальном рассмотрении каждого года обнаруживаются существенные температурные различия. Так, за период с 2003 по 2016 гг., (значения температуры поверхности получены по данным GHRSST Level 4 MUR Global Foundation SST Analysis (v4.1), а также – MODIS Aqua (SST)) наиболее теплыми годами являлись 2010, 2005 и 2004 гг. (среднегодовые значения, соответственно, – 19,287, 19,179 и 19,083°С), а самыми холодными – 2009, 2008 и 2003 гг. (среднегодовые значения, соответственно, – 17,864, 18,060 и 18,177°С). Экстремальные цветения сине-зелёных водорослей наблюдались как в теплые годы (2005 и 2010 гг.), так и в холодные (2008 и 2009 г.). При рассмотрении исключительно поры «гидрологического лета» (май-сентябрь) наблюдалось, что фаза начального развития экстремального цветения сине-зелёных водорослей начиналась при температурах поверхности от 26,5 до 30°С и более, а фаза максимального развития – в диапазоне (по осредненным данным) от 26,1 до 29,3°С. Наиболее длительное и интенсивное цветение регистрировалось в 2005 г с 5-7 августа по 26-27 сентября. Фаза начального развития длилась порядка 2,5 недель, видовой состав фитопланктона по данным дистанционного зондирования не идентифицировался, в подповерхностном слое в центральной части Южного Каспия формировалась структура, которая постепенно увеличивалась в масштабах. С 24 августа на оптических изображениях среднего разрешения по составленным RGB-композитам, а также по изменениям значений яркости восходящего излучения определяется наличие фитопланктона на поверхности – из-за свойства положительной плавучести цианобактерии проявляются не только в видимом диапазоне (469-645 нм), но и частично в ближнем инфракрасном (859 нм). Фаза максимального развития длится до 17 сентября, во время которой площадь захваченных аномальным цветением вод превышает 35000 км² (на 1 сентября), а максимальная протяженность достигает 388 км по условной линии «Ленкорань – Горган» (рис. 2). С 17 сентября наступает завершающая фаза – фаза разрушения, длящаяся до 26-27 сентября.
Возникновение и формирование аномального цветения происходит над глубоководной частью Южного Каспия, хотя высокое количество фитопланктона ранее обнаруживается практически вдоль всего южного сектора береговой линии. В дальнейшем, по мере эволюции, наблюдается некоторая пространственная миграция: увеличиваясь в размерах, цветение захватывает воды сначала юго-восточнее, а затем еще и северо-западнее. На пике максимального развития экстремальное цветение сначала по площади достигает максимальных величин, затем несколько сокращается в размерах, также продолжается миграция: область цветения смещается на юго-запад и достигает берега, где и прекращает свое существование в конце регрессивной фазы. На протяжении всех фаз эволюции сине-зелёные водоросли служат хорошим индикатором динамической активности бассейна: прослеживается влияние вихревых структур в начале развития, отчетливо выделяются вихри на протяжении стадии максимального развития цветения. В высоком разрешении (рис. 3) также явно различимы процессы мелкомасштабного и субмезомасштабного уровней.
Цветения сине-зелёных водорослей в 2008 и 2009 гг. отличались меньшими масштабами и меньшей продолжительностью, чем в 2005 г. Детальный анализ этих ситуаций затруднен из-за большого количества дней с развитым облачным покровом над южной частью Каспийского моря. В 2008 г. аномалия начинает формироваться в августе несколько южнее центра глубоководной части, и постепенно, переходя к стадии максимального развития, смещается на запад. Жизнедеятельность сине-зелёных водорослей на поверхности определяется непродолжительное время (с 26 по 29 сентября), а к 3-5 сентября аномалия разрушается, и цианобактерии заглубляются. Аномальное цветение 2009 г. так же, как и в 2008 г., затруднено для детального анализа из-за нередкого полного или частичного закрытия района облаками. В целом, по спутниковым данным, первые признаки цветения (увеличение количества биомассы фитопланктона в толще вод) обнаруживаются в начале августа над глубоководной котловиной Южного Каспия. По мере развития аномалии и миграции сине-зелёных водорослей к поверхности происходит смещение цветения на юго-запад к прибрежной зоне. С 18-20 по 22-23 августа на смену начальной фазе приходит фаза максимального развития: аномальное цветение по значениям отраженной компоненты определяется как находящееся на поверхности. Затем, при переходе в стадию регрессии, цветение сдвигается в северном направлении, сама аномалия прекращает свое существование 5-6 сентября. В 2010 г. экстремальное увеличение биомассы сине-зелёных водорослей наблюдалось дважды: фазы максимального развития регистрировались в июле (продолжительность составила 3-5 дней) и в августе (длительность – 3 дня). Первые признаки формирования аномального цветения обнаруживаются в первой трети июля в юго-восточной части Южного Каспия, с 10-12 июля фиксируется повышенное количество биомассы фитопланктона. Аномалия под влиянием динамических процессов в некоторой степени меняет структуру, но в целом сохраняет свое местоположение: юго-восточный район и вдоль восточного берега на границе шельфа. С 21 июля уверенно определяется наличие сине-зелёных водорослей на поверхности: структура аномалии имеет нитевидную форму, ориентированную на северо-запад. Часть структуры, находящаяся в подповерхностном слое, продолжает развиваться уже над глубоководными районами и достигает западной половины акватории. Скопление цианобактерий на поверхности сохраняется непродолжительное время, и уже с 23-24 июля повышенное содержание фитопланктона обнаруживается только в толще вод. Такая ситуация, при которой наблюдается невыраженная, но сложная миграция аномалии (к этому времени захвачено около двух третей вод преимущественно в юго-восточной половине акватории), продолжается до 1 августа, а со 2 августа регистрируется наличие цианобактерий на поверхности в юго-западном районе на значительно расстоянии от берега. В эти же дни (2-3 августа) выход на поверхность сине-зелёных водорослей происходит также вблизи южного берега. Оба фрагмента существуют до 9 августа, а с 10 августа и до последних чисел месяца наблюдается только повышенное содержание фитопланктона в толще вод, развивающееся в центральной части Южного Каспия. Причем, высокая концентрация биомассы водорослей фиксируется до 21 августа, в дальнейшие же дни количество фитопланктона значительно снижается.
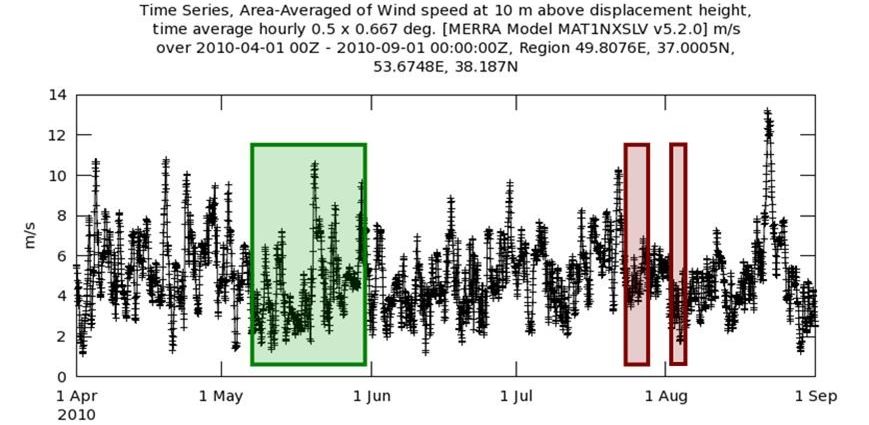
На протяжении 2011-2016 гг. аномальное цветение в южной части моря не возникало (за исключением локальных случаев в прибрежных зонах, не определяемых по данным дистанционного зондирования), и следующая серия вспышек была обнаружена только в 2017 г. В отличие от всех ранее рассматриваемых лет, летом 2017 г цветение сине-зелёных водорослей формируется во всем Каспийском море: в северной части (что типично), в южной и впервые – в средней (рис. 4). Повышенное содержание биомассы фитопланктона выявляется 19-20 июня в Среднем Каспии северо-восточнее Апшеронского полуострова. В последующие дни наблюдается рост количества водорослей в подповерхностном слое, и 27 июня насыщение вод фитопланктоном хорошо различается в видимом спектре: цветение захватывает полностью воды Северного и Среднего Каспия, и частично – Южного. Тем не менее, цветение сине-зелёных водорослей с развитием на поверхности регистрируется только спустя месяц – 27-28 июля в юго-восточной части Южного Каспия (аномалия имеет вытянутую структуру с грибовидным окончанием, ориентированную от берега на юго-юго-запад). В дальнейшем (с 31 июля и первых чисел августа) происходит серия возникновения аномалий и в Южном Каспии, и в Среднем, с максимумом развития в пределах 4-5 – 9 августа (по числу фрагментов), а также – 11-14 августа (меньшее число фрагментов вдоль западной береговой линии и южной в средней и южной частях бассейна). С 5 по 7 августа (на пике развития) можно выделить до 6 аномальных участков (по критерию нахождения сине-зелёных водорослей на поверхности), которые, скорее, являются фрагментами двух обширных зон экстремальной концентрации цианобактерий: в западной половине Среднего Каспия с проникновением в Южный Каспий и отдельно – в южной половине Южного Каспия. Вторая «волна» цветения менее выражена: 11 августа сохраняются фрагменты аномалии на юге моря (в фазе регрессии), возникают новые в средней части моря вблизи западного берега, которые фиксируются до 15 августа. С 16 августа повышенное содержание фитопланктона наблюдается лишь в толще вод. Неизбежно возникает вопрос, какие условия способствуют возникновению и дальнейшему развитию аномальных цветений как типичного для моря фитопланктона, так и сине-зелёных водорослей. Ранее указывалось, что определенная роль отводится температуре: экстремальное увеличение биомассы фитопланктона (как типичного, так и цианобактерий) возможно при достаточном прогреве вод. Но увеличение температуры деятельного слоя – не единственное условие: для эволюции аномального цветения также имеют значение интенсивность перемешивания поверхностного слоя вод и активность поступления биогенных элементов. Над южной частью Каспийского моря в летнее время преобладают ветра невысоких скоростей, в среднем не превышающие 5-6 м/с. Но при этом от года к году в отдельные месяцы наблюдаются существенные вариации синоптической обстановки: регистрируются как периоды ветрового затишья (скорости ветра невелики и устойчивы), так и периоды, когда ветер характеризуется неустойчивым направлением и скоростями, достигающими 8-10 м/с и выше. При оценке ветрового поля непосредственно перед началом возникновения того или иного вида цветения, а также на протяжении его существования отмечается две характерные ситуации: аномальному цветению типичного фитопланктона предшествует период с высокими и неустойчивыми скоростями ветра, а аномальное цветение сине-зелёных водорослей (особенно – максимальная фаза эволюции с развитием цианобактерий на поверхности) происходит после или на фоне малых и устойчивых скоростей ветра. В той или иной степени это соответствие выявляется во всех случаях аномального цветения, в отдельные годы оно выражено явно. Например, в 2010 г существовало два типа цветения: аномальное развитие характерного для акватории фитопланктона в мае и интенсивный рост сине-зелёных водорослей в июле и августе. В весенний период ветру свойственны высокие скорости (до 8-10 м/с), перемежающиеся за малые временные периоды с низкими (3-6 м/с). Интенсивное ветровое воздействие обеспечивает активное перемешивание поверхностного слоя моря, что усиливает газообмен на границе «вода-атмосфера» и способствует развитию фитопланктона. Иная синоптическая обстановка соответствует периодам максимума цветения сине-зелёных водорослей: пик их развития совпадает с днями, когда скорости ветра были малы (3-7 м/с) и устойчивы (рис. 5). При таких ветрах волнение на поверхности моря развито слабо, перемешивание вод ослаблено, что позволяет сине-зелёным водорослям, обладающим свойством положительной плавучести, формировать колонии на нулевом горизонте (тогда как при интенсивном ветровом перемешивании колонии заглубляются и разрушаются).
Экстремальное цветение наблюдается также в случаях, когда оптимальные ветровые условия наблюдались не во время фазы максимального развития, но на протяжении длительного времени до возникновения цветения или же во время начальной фазы. Так, в 2001 г регулярные подъемы скорости ветра до 8-12 м/с наблюдались с мая до середины июля (рис. 6), то есть, на протяжении длительного времени происходило активное перемешивание поверхностных вод. А когда поверхность моря значительно прогрелась (примерно до 28,5-29°С), начался рост биомассы типичного фитопланктона.
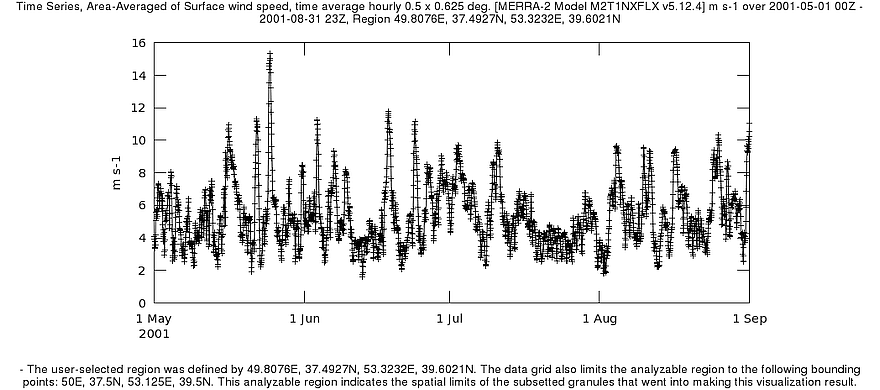
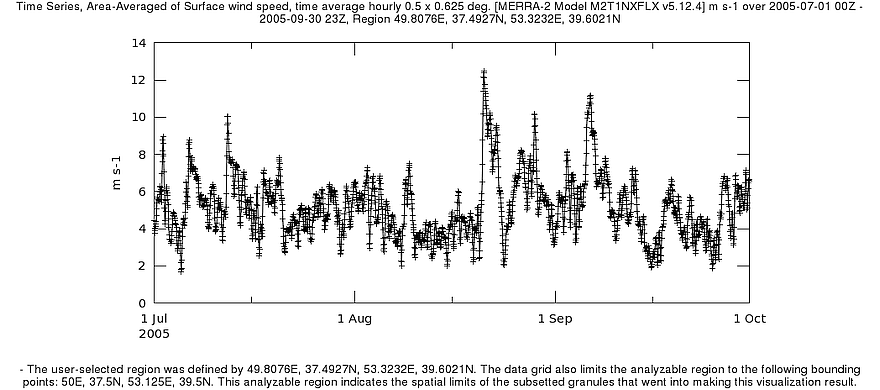
В 2005 г, когда было зарегистрировано обширное цветение сине-зелёных водорослей, летом наблюдалось длительное ветровое затишье: 16 июля по 19 августа над южнокаспийской акваторией преобладал ветер с достаточно слабыми и мало изменчивыми скоростями – от 3 до 5,5 м/с (рис. 7). В этот период возникла и стала формироваться аномалия (начальный этап). К тому моменту, когда характер ветра изменился (снизилась устойчивость, появились скачки до 8-12 м/с, периоды затишья стали кратковременными), количество биомассы цианобактерий достигло столь существенных значений, что развитие продолжилось на фоне сравнительно неблагоприятных условий.
Несмотря на немаловажное значение гидрометеорологических условий для возникновения экстремальных цветений, ключевая роль принадлежит биогенным элементам. Для акватории Южного Каспия основными источниками биогенных элементов могут быть реки и частые для данного региона пыльные бури. Хотя реки, впадающие в южную часть моря, маловодны, косвенно на их влияние указывает то, что регулярно происходит заметное увеличение биомассы фитопланктона в сравнительной близости от приустьевых областей. В дальнейшем часть этих формирований (распространяемых, в том числе, в глубоководные районы) становится основой для эволюции аномального цветения (табл. 2). Наиболее существенное значение для южнокаспийского бассейна оказывают реки Кура, Сефидруд, Горган и Атрек. Все перечисленные реки активно используются в хозяйственной деятельности и испытывают значительную антропогенную нагрузку [1, 17, 24].
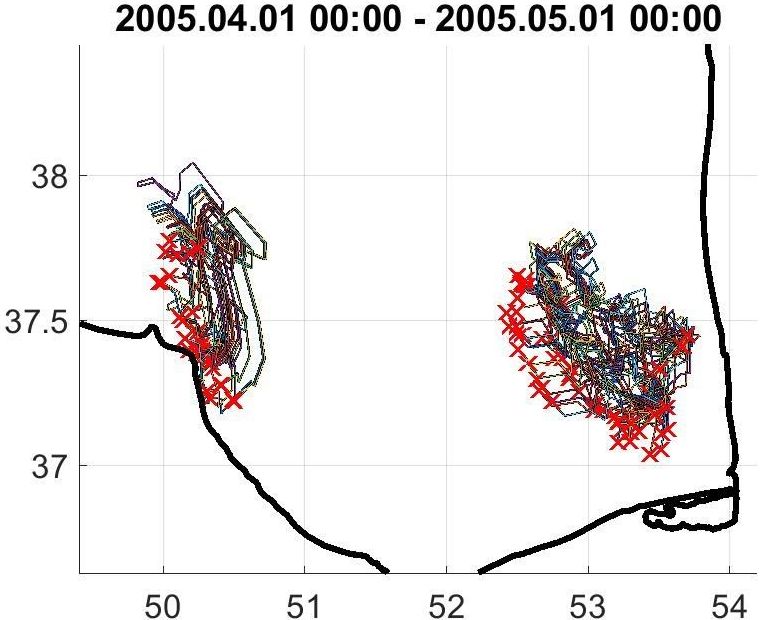
При выделении речных плюмов обозначенных рек на спутниковых снимках обычно наблюдается транспорт взвесей динамическими процессами вдоль береговой линии, но в ряде случаев они простираются на более значительные расстояния от устья. Для получения дополнительных данных о распространении пресных вод производились вычисления с помощью программы FOTS, моделирующей перенос природных объектов на основе данных спутниковой альтиметрии и данных о полях скорости ветра (высота 10 м). В силу того, что у берега (0-30 м) альтиметрические данные имеют значительную погрешность, точки, обозначающие источники речных вод, выставлялись на некотором отдалении от устьев: одна вблизи дельты Сефидруда, две других – в районе выноса вод Горгана и Атрека. Время задавалось с 1 по 30 апреля с учетом того, что в этот период реки наиболее полноводны, и их сток – максимален. В итоге воспроизводились траектории плавающих объектов (пресных вод), которые отображали их перенос по акватории Южного Каспия (рис. 8).
При совмещении спутниковых снимков с результатами моделирования (по географическим координатам), выявилось, что формирование аномальных цветений сине-зелёных водорослей происходило в районах влияния южнокаспийских рек (рис. 9).
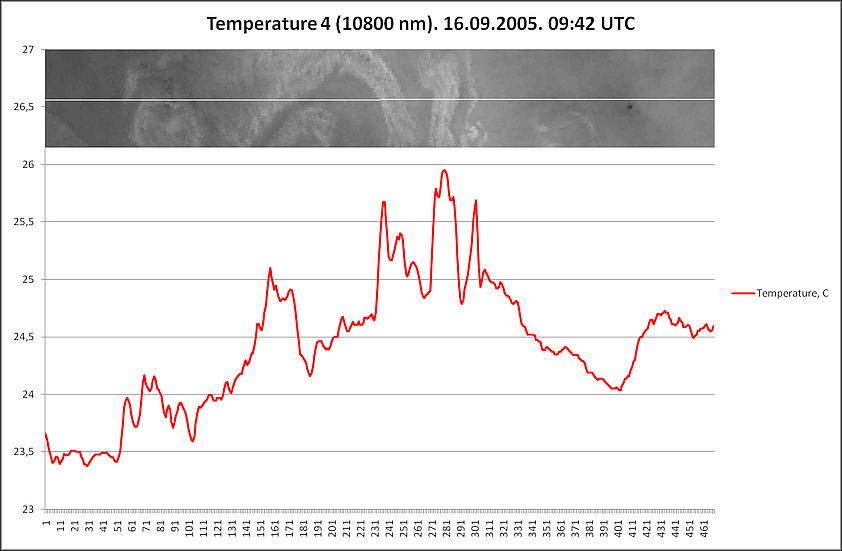
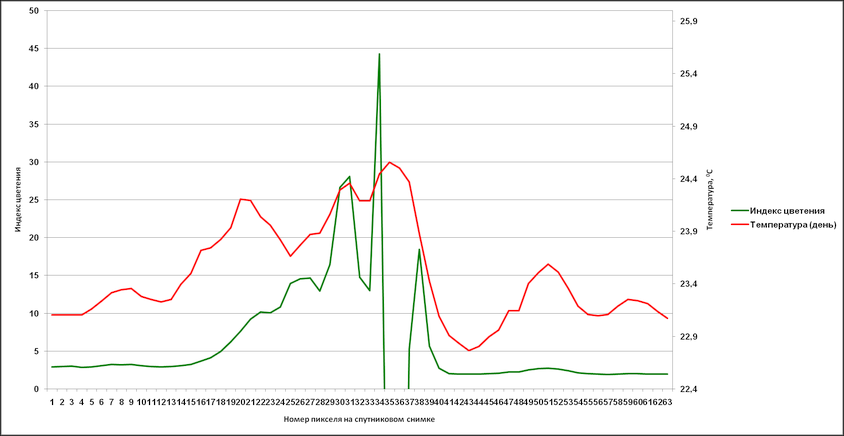
Стоит отметить, что в летний период подобные совпадения практически не обнаруживаются, и речные воды по результатам расчетов распространяются на расстояниях меньшие от берега, чем возникшие очаги цветений. Тем не менее, допустимо рассматривать реки в качестве источника биогенных элементов с учетом того, что насыщение вод питательными элементами могло происходить заведомо раньше возникновения аномального цветения – до ситуации оптимального гидрометеорологического фона. Определённый вклад по занесению биогенных элементов Каспийское море может принадлежать также и пыльным бурям, хотя прямая связь между возникновением аномальных цветений (сине-зелёных водорослей) и интенсивностью пыльных бурь не обнаруживается. Но замечено, что наиболее часто в летние месяцы пыльные бури проходят над Южным Каспием (до 7-15 дней в месяц), тогда как над Средним Каспием и Северным данные явления фиксируются реже (до 3-9 дней в летний месяц и от 0 до 2-3 дней соответственно). Схожая ситуация выявляется при оценке количества почвенных и пылевых частиц над акваторией Каспийского моря. В среднем, за период с января 2003 по декабрь 2017 г. над Северным Каспием улавливается от 1,455х10-11 до 2,660х10-11 кг/м²·с, над Средним – от 1,588х10-11 до 2,794х10-11 кг/м²·с, над Южным – от 1,978х10-11 до 3,257х10-11 кг/м²·с. При комплексном воздействии различных факторов создается фон, благоприятный для развития того или иного вида аномального цветения. Но и само цветение, когда биомасса фитопланктона достигает экстремальных значений, оказывает влияние на окружающую среду. Так, выраженное цветение сине-зелёных водорослей за счет свойства положительной плавучести создает теневые зоны, которые угнетают развитие иных видов фитопланктона, находящихся в нижележащих слоях. Также цианобактерии при массовом своем размножении создают в акваториях эффект вторичного загрязнения, который, в свою очередь, увеличивает риск эвтрофикации вод. И, конечно, особую опасность представляют собой выделяемые в воду токсины, что при высокой концентрации приводят к заморным явлениям, становятся причиной отравления и гибели птиц и млекопитающих. С помощью данных дистанционного зондирования было дополнительно обнаружено влияние аномального цветения сине-зелёных водорослей на физические характеристики зараженных вод: их температура в отдельные дни существенно превышала температуру вод, в которых цианобактерии по спутниковым снимкам не обнаруживались. Для более детального исследования была использована ситуация с аномальным цветением в 2005 г (наиболее обширное и продолжительное цветение за весь период исследования). Был отобран ряд данных NOAA-17 с учетом особенностей изображений в видимом диапазоне: отсутствие облачности или незначительное количество облаков, полный охват снимком области в пределах 48° з. д. до 55,3° (55°29’33”) з. д. Эти координаты обозначили проложенный вдоль 38° с. ш. профиль, что пересекал аномалию цветения и частично захватывал свободные от цианобактерий воды. Анализировались все каналы обозначенных спутниковых данных (яркость восходящего излучения и температура поверхностного слоя), наибольшее внимание уделялось оптическому каналу в красном спектре (длина волны – 630 нм) и инфракрасному каналу (длина волны – 10800 нм). С учетом качества снимков было выбрано несколько дат, отражающих изменения аномального цветения и его влияния на водную среду: 15 августа, 20 августа, 10 и 16 сентября. В середине августа (15 числа), цветение сине-зелёных водорослей пребывает в фазе начального развития, водоросли на поверхности не фиксируются, площадь захваченных вод достигает 9586 км². Зона высокой концентрации фитопланктона хорошо различима на канале с длиной волны 630 нм, частично – 3740 и 10800 нм. Температурные различия между областью аномалии и чистыми водами практически не выявляются. К 20 августа площадь аномального цветения увеличивается до 24651 км², сине-зелёные водоросли по-прежнему обнаруживаются только в толще вод. Как и 15 августа, цветение проявляется на длинах волн 630 нм (отчетливо), 3740 и 10800 нм (частично). Температурные отклонения присутствуют, но не позволяют указать на прямую связь с интенсивностью развития сине-зелёных водорослей. Иная картина наблюдается в сентябре, когда цветение находится в фазе максимального развития. На 10 сентября площадь захваченных вод несколько меньше максимально зарегистрированной (35000 км² в начале сентября) – около 29811 км², цианобактерии образуют плотные скопления не только в толще вод, но и на поверхности. Аномалия четко выделяется в диапазоне от 630 до 12000 нм данных спутника NOAA-17, также хорошо различима при создании изображения (рис. 10).
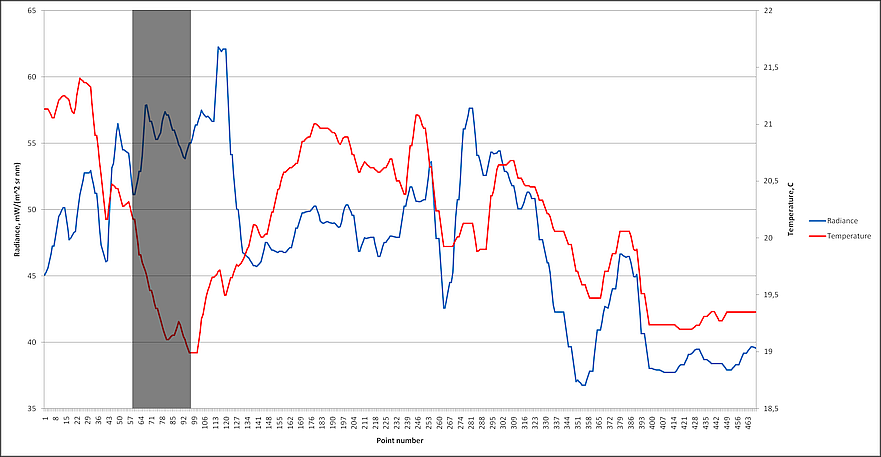
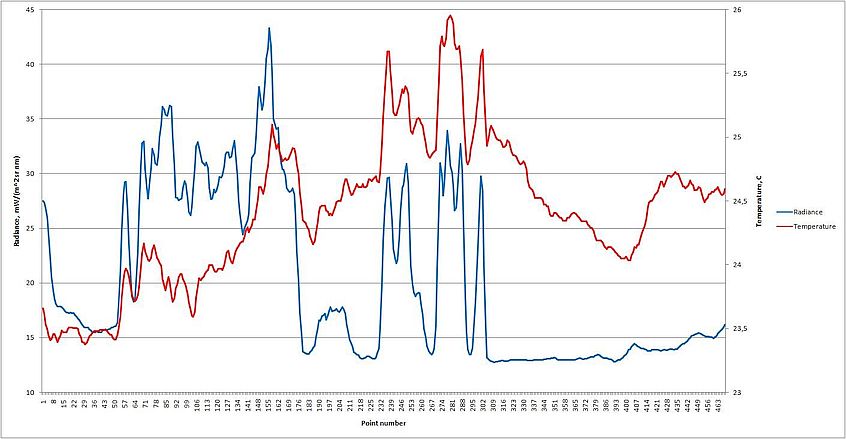
Различия между зараженными водами и свободными от цветения отображаются в данных (по линии профиля) яркости восходящего излучения и температуры поверхности, при их сопоставлении фрагментарно прослеживается положительная корреляция, когда изменения значений яркости восходящего излучения вызывают соответствующий отклик температуры (рис 11). Максимальная разница температуры в районе аномалии и свободных вод превышает 2°С.
К 16 сентября количество биомассы сине-зелёных водорослей несколько снижается, цветение находится в стадии перехода в регрессивную фазу, хотя площадь аномалии по-прежнему велика (25137 км²), и сохраняется положение цианобактерий на поверхности. Цветение, как и 10 сентября, обнаруживается на всех каналах в пределах 630-12000 нм. На графиках, полученных по данным, соответствующим линии профиля, экстремальное цветение отражается и в изменениях значений яркости восходящего значения, и колебаниях температуры поверхности (рис. 12). При их (излучение и температура) сопоставлении наблюдается выраженная связь, прогрев зараженных вод достигает 2,5°С (рис 13).
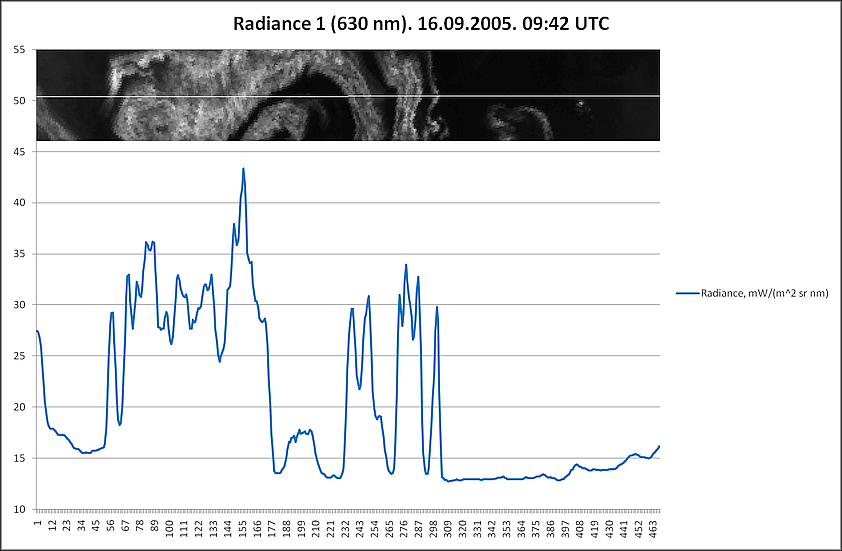
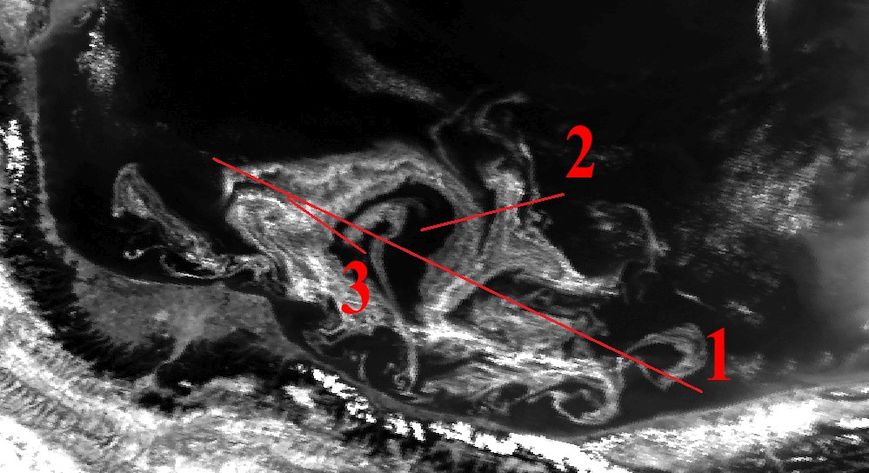
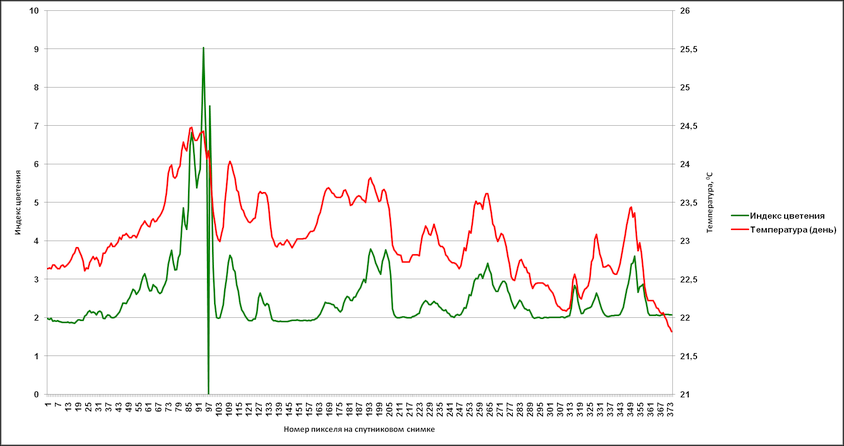
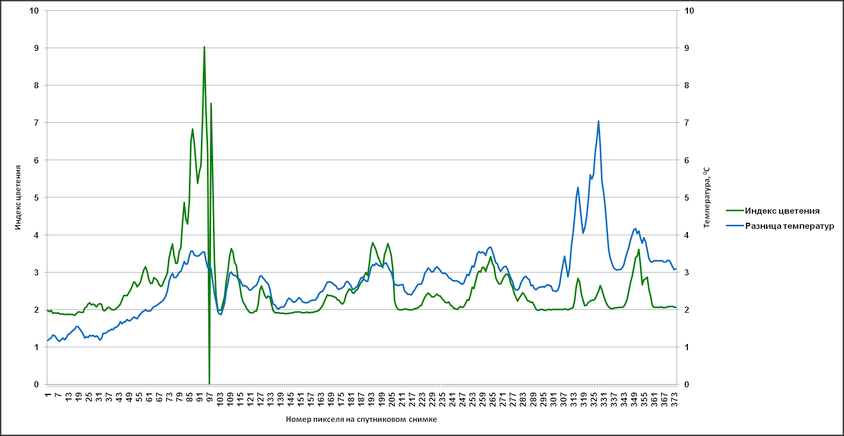
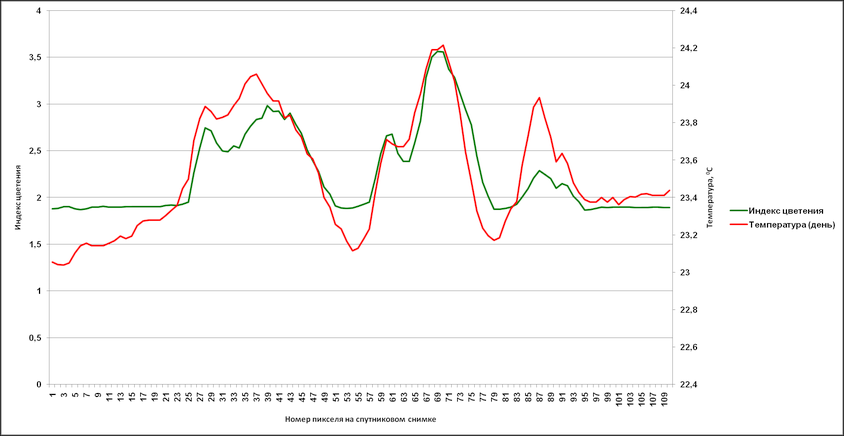
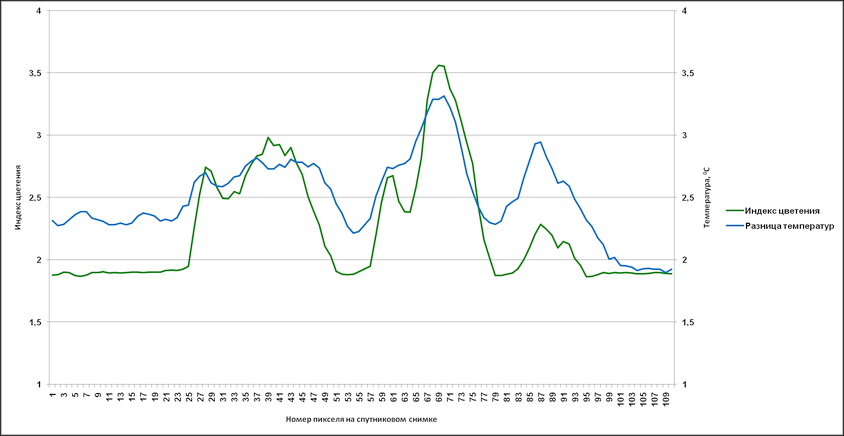
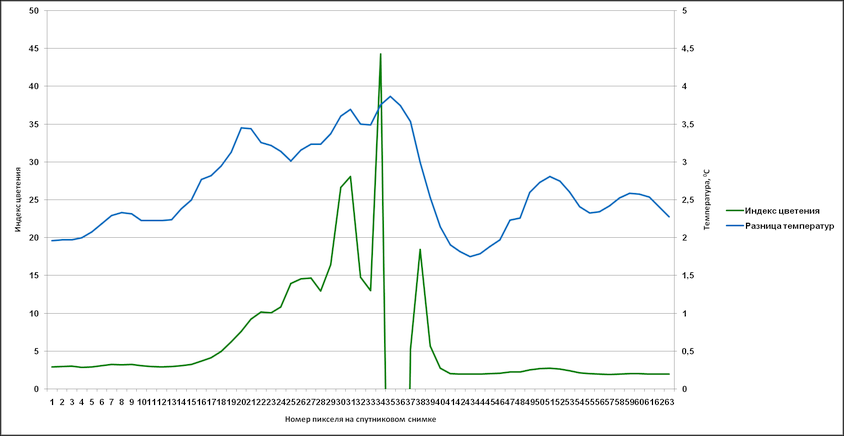
Аналогичные результаты были получены и при привлечении индекса цветения, полученного путем комбинации каналов с длинами волн 630 (R1) и 862,5 nm (R2) – (R1-R2/R1+R2). Были сгенерированы профили, пролегающие через различные участки аномального цветения там, где на снимке отсутствует или минимален облачный покров (рис. 14).
На основании данных, соответствующих этим профилям, в дальнейшем были созданы графики с отражением изменений индекса цветения и температуры поверхности (дневная, разница между дневной и ночной) (рис. 15-17). Индекс цветения реагирует на количество биомассы фитопланктона, в зонах наиболее высокого содержания сине-зелёных водорослей он достигает значений 40-45 единиц. Также воспроизводятся температурные отклонения: в тех случаях, когда профиль пролегает и через воды, насыщенные цианобактериями, и воды, свободные от них, контраст температур достигает 1,5-2,5°С (а на протяжении фазы максимального развития выявлялся перегрев на 3°С и более). Наиболее высокое соответствие изменений температуры и индекса цветения выявляется в данных профиля 2 – положительная корреляция составляет 0,88 (рис. 16а) и 0,84 (рис. 16б). Профиль 3 нанесен на спутниковый снимок таким образом, что пересекает облачный покров, что отражается на графиках в виде отрицательных значений индекса цветения.
Заключение. В Каспийском море аномальные цветения фитопланктона, которые представляют собой резкое увеличение количества фитопланктона на единицу объема воды, регистрируются ежегодно в северной части, крайне редко – в средней и в отдельные годы – в южной части. В Южном Каспии, в отличие от Северного, вклад речного стока невелик, и поэтому возникновение и развитие аномальных цветений обеспечивается комплексным влиянием различных факторов. По данным дистанционного зондирования за период с 2001 по 2017 гг. аномальное цветение фитопланктона в южной части Каспийского моря было выявлено в 2001, 2005, 2008, 2009, 2010 и 2017 гг. в летний (вторая половина лета) или летне-осенний периоды. В 2001 и 2010 гг. видовой состав цветения определял характерный для акватории фитопланктон, тогда как в остальные годы (включая и 2010 г) аномалии формировались из-за экстремального увеличения количества сине-зелёных водорослей (преимущественно рода Nodularia Spumigena). Периоды цветения цианобактерий отличались различной продолжительностью (от дней до нескольких недель) и условно делились на три фазы: начального развития, максимального развития и исчезновения. Для характеристики аномалий цветения наибольший интерес представляет фаза максимального развития, когда сине-зелёные водоросли достигают поверхности и уверенно распознаются по характерным признакам на спутниковых изображениях оптического диапазона. В течение периода существования аномальное цветение фитопланктона может мигрировать под влиянием динамических процессов на значительные расстояния, на пике развития – захватывать водные пространства площадью в сотни и тысячи квадратных километров. В южной части Каспийского моря экстремальное цветение формируется на фоне ряда факторов – гидрометеорологических условий и поступления биогенных элементов. В частности, увеличение биомассы фитопланктона происходит при значительном прогреве деятельного слоя вод (до 26,5°С и выше) и либо интенсивном ветровом воздействии (возможен рост биомассы типичного фитопланктона), либо, наоборот, при устойчивых скоростях ветра не более 5-7 м/с (тогда создаются оптимальные условия для возникновения выраженного цветения сине-зелёных водорослей). В процессе исследования на спутниковых снимках, отражающих температуру поверхности моря, обнаружились зоны перегрева, совпадающие по времени и местоположению с существующими аномалиями цветения сине-зелёных водорослей. При более детальном изучении вопроса выявилось, что воды в зонах аномального цветения имеют более высокую температуру, чем воды, свободные от цветения – температурный контраст зависит от степени развития аномалии и может достигать 1-3°С. Литература:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||